The Organisation and Growth of Plant Bodies in Comparison with Animals

Figure 2 Diagram of a Plant Cell
Click thumbnail to view full-size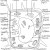
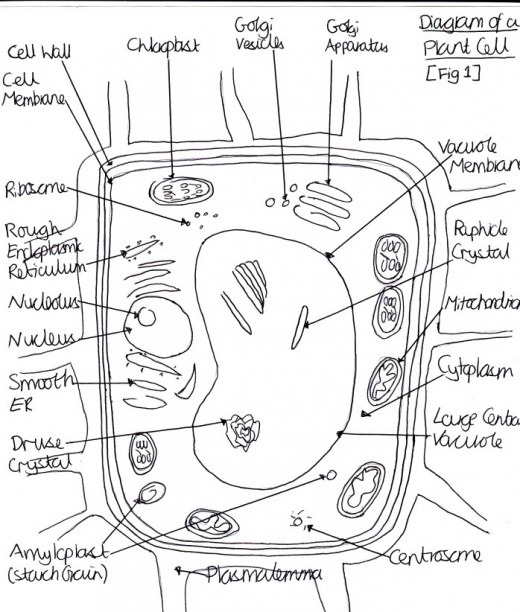
Figure 1 Diagram of a True Leaf
Click thumbnail to view full-size
Organisation of Plant Bodies
Plant cells are formed at meristems, and then develop into cell types which are grouped into tissues (Matthews, 1999) [Figure two].
The tissues in the plant include [Figure one]: the dermal tissue (which cover the outer surface of herbaceous plants and is composed of closely packed epidermal cells), ground tissue (which are the bulk of the primary plant body, e.g. parenchyma, collenchyma, and sclerenchyma cells) and vascular tissue (which transports food, water and nutrients around the plant e.g. xylem, phloem, parenchyma and cambium cells) (Alberts, 2002) [Table 1].
The primary plant body is the extension of the root and stems.
The shoot system is made up of the stems and their leaves, which includes: apical meristems (regional localised mitosis that causes primary growth at the shoot tips and root tips), axillary buds (lateral shoot apexs that allow replacement of the main shoot), internodes (the spaces between leaf attachments) and organs (e.g. leaves, stem, flowers and fruit) (Alberts, 2002).
The root system is what anchors the plant, and absorbs water and nutrients. It consists of: apical meristems, and organs (e.g. roots, tubers, rhizomes).
Table 1 Major Tissues, Cells and Cell Types
Major Tissue
| Cells and Cell Types
|
|---|---|
Dermal
| Epidermal-trichrome, subsidiary, epidermal and guard cell
|
Peridermal-phellem, phellogen and phelloderm
| |
Vascular
| Xylem-vessel member, tracheid, fibre and parenchyma cells
|
Phloem-sieve tube members, sieve cells, companion cells, fibre, sclerid and parenchyma cells
| |
Cambium
| |
Ground
| Parenchyma
|
Collenchyma
|
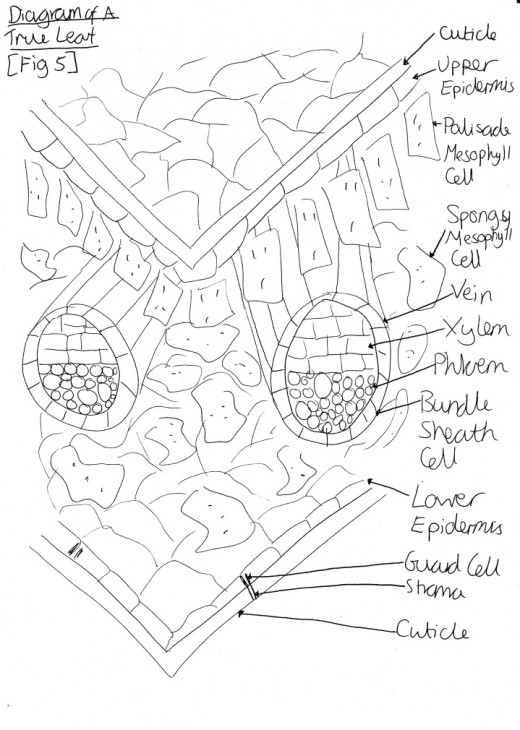
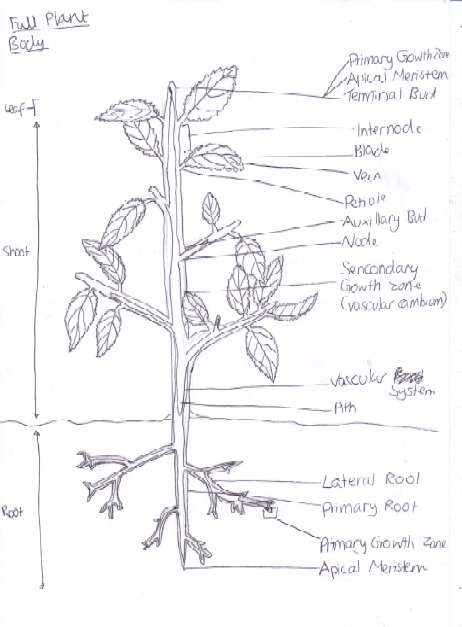
Figure 7 Diagram of a Full Plant Body
Click thumbnail to view full-size
Growth of Plant Bodies
Plants grow from their meristems, which is a region of localised mitosis. Meristem cells have a potentially unlimited ability to divide, however this is controlled. One type of meristem is the lateral meristems for example branches. They are cylinders that extend almost the whole length of the plant. For example, monocots have intercalary meristems that are located between differentiated tissues at the base of each internode (Alberts, 2002). Another meristem type is cambium, e.g. xylem and phloem. It is also lateral but is able to produce secondary growth and so can produce wood or cork (The Forestry Commission, 2012). One further meristem type is the apical meristems, e.g. shoot or root tips [Figure seven].
Apical dominance is when the intercalary (lateral) meristems are suppressed while the tip is able to continue growing [Figure seven]. This allows the nutrients of the plant to be focused on one area for whatever function the plant is trying to achieve e.g. whether it is to grow taller for more light or whether it needs to concentrate on reproduction. This process is able to happen because information is exchanged between the meristems (Alberts, 2002).
Growth of Animal Bodies
Animal growth, however, is different. Instead of the idea of apical dominance, animals have allometric growth, thus meaning that all their body parts grow uniformly so the overall proportions are maintained. If plants were to do this, it would be less efficient for them to photosynthesise because the stem cannot capture the sun’s energy and so it is extra live tissues which need to be provided with nutrients and water (Alberts, 2002). Another difference between plants and animals is that the meristem calls in plants have the potential to divide infinitely however animals stop growing after they have reached their full size. Also, plants are able to grow new organs, which is used in horticulture (e.g. Begonia spp. leaf cuttings to grow entirely new plants) but they are unable to repair old or damaged cells and tissues. However, animal cells can divide to repair but cannot create new organs e.g. epithelial cells and blood stem cells. Gymnosperms and dicots have extensive lateral meristems (e.g. cambium, vascular cambium and cork cambium) that cause lateral growth but only at certain times of the year, which creates annual growth rings-However, animals do not have annual growth (Alberts, 2002).
References
Alberts, B. et al. (2002), Molecular Biology of the Cell [4th edition], Garland Science.
Boerjan, W. (2010), ‘Initial Field Test Results GM Poplars: Bioethanol Yield Almost Doubled’ [online journal], Available at http://www.vib.be/en/news/Pages/Initial-field-test-results-GM-poplars-bioethanol-yield-almost-doubled.aspx, (Accessed: 24/11/12).
Cossalter, C. et al. (2003), Fast-Wood Forestry: Myths and Realities, Indonesia Printer, Available at http://www.cifor.org/publications/pdf_files/Books/ForestPerspective.pdf, (Accessed: 24/11/12).
‘Frontier in Plant Physiology’ (page 1) [online journal], Available at http://www.frontiersin.org/Plant_Physiology/10.3389/fpls.2012.00085/abstract, (Accessed: 24/11/12).
Guardian, ‘The GM Tree Plantation Bred to Satisfy the World’s Energy Needs’, Available at http://environment/2012/nov/15/gm-trees-bred-world-energy, (Accessed: 24/11/12).
Lybeer, B. (2006), ‘Lignification and Cell Wall Thickening in Nodes of Phyllostachys viridiglaucescens and Phyllostachys nigra’, Annuals of Botany, Volume 97 (pages 529-539), [online journal], Available at http://aob.oxfordjournals.org/content/97/4/529.full.pdf, (Accessed: 20/11/12).
Matthews, C. et al. (1999), Biochemistry, [3rd edition], Benjamin Cummings.
Meike (2012), ‘Inexpensive, Environmentally Friendly and High Permeable Lignin-Based Ion Exchangers’, Industrial Partnership Office, Available at https://ipo.llnl.gov/?q=technologies-lignin_based_ion_exchangers, (Accessed: 24/11/12).
Pauly et al. (2007), ‘Cell Wall Carbohydrates and their Modification as a Resource
for Biofuels’, The Plant Journal, Volume 54 (page 1) [online journal], Available at http://public.wsu.edu/~lange-m/Documnets/Teaching2011/Pauly2008.pdf, (Accessed: 24/11/12).
Pollack, A. (2007), ‘Through Genetics, Tapping a Tree’s Potential as a Source of Energy’, The New York Times, Available at http://www.nytimes.com/2007/11/20/science/20tree.html?pagewanted=all, (Accessed: 24/11/12).
Scheller H., Ulvskov, P. (2010), ‘Hemicelluloses, Annual Review of Plant Biology’, Volume 61 (page 1), [online journal], Available at http://www.annualreviews.org/doi/abs/10.1146/annurev-arplant-042809-112315, (Accessed: 22/11/12).
Strnad (2000), ‘Cellulose Fibres Functionalised by Chitosan: Characterization and Application’ [online journal], Available at http://cdn.intechopen.com/pdfs/12146/InTech-Cellulose_fibres_funcionalised_by_chitosan_characterization_and_application.pdf, (Accessed: 24/11/12).
The Forestry Commission (2012), ‘Replace Other Materials with Wood’, Available at http://www.forestry.gov.uk/forestry/INFD-7M8MXV, (Accessed: 22/11/12).
Underwood, W. (2012), The Plant Cell Wall: A Dynamic Barrier Against Pathogen Invasion, UXL Encyclopaedia of Science (2002), (Second edition), The Gale Group Inc.
Vidal, J. (2012), The GM Tree Plantations Bred to Satisfy the World's Energy Needs.
Zaikov, G. et al. (2004), Polymer Analysis, Degradation and Stabilization (Pages 97-100), Nova Science Publishers Inc.
 Plant With The Most Benefits")

